

In our quest to engineer human levels of performance in robots, I and others in robotics have been trying to imitate the macroscopic behavior of muscle. We have superficially simulated the springlike properties of muscles and their role in control using force controlled electric motors and hydraulics. We have followed a design path of ever higher performance force and torque sensing and lower latency velocity and torque feedback control, resulting in ever more expensive robots. Perhaps we have focused on the wrong aspect of muscle. This review focuses on muscle damping, and how imitating it may help us build less expensive robots with a more human-like touch
We are attempting to bridge muscle molecular mechanisms and macroscopic behavior. A key question is "How is muscle so efficient when acting as a brake?" When we compare muscle to robot actuators, it is also much more efficient. Motors used in robots use a lot of energy to hold a position or resist a push. Electric motors produce torque that is proportional to current, and the power used is the current multiplied by the power supply voltage. Hydraulic and pneumatic motors can hold a position with no energy cost by closing valves. However, resisting a pull while moving has an energy cost of the fluid flow multiplied by the supply pressure. In addition, force-controlled hydraulic systems used on humanoid robots typically have significant continual energy loss through internal leakage, which is necessary to keep friction in pistons low. To eliminate these costs for holding a position or resisting a pull, typically friction brakes need to be added at each joint.
How does muscle not burn a lot of energy under load when the muscle (not including the tendon) is staying at the same length (isometric) or lengthening under load (doing negative work)? The crossbridges are bathed in stored energy (Adenosine Triphosphate, ATP) all the time, so it is not that the energy source is removed. If ATP hydrolysis is used for crossbridge detachment, why doesn't any form of crossbridge cycling (which has to happen in muscle lengthening under load) burn energy?

The next slide describes where the talk is going.
It has been recently confirmed that skeletal muscle myosin shares with other myosin isoforms a catch-bond property. The myosin head is much less likely to detach from the actin filament if the crossbridge is stretched. In addition, it has been hypothesized that there are rapid crossbridge detachment and reattachment processes that do not require the use of stored energy in the form of Adenosine Triphosphate (ATP). These processes allow a crossbridge to translate or "jump" along the actin filament to continue to resist a muscle being lengthened. The catch-bond and jump properties allow muscle to macroscopically act as a brake with no use of energy when lengthening, as well as like a spring for small perturbations. These properties also imply that large scale use of ATP only occurs when the muscle is shortening, and only when the muscle is doing positive work as a unidirectional motor. The nervous system can move the muscle to the motor mode with sufficient activation to start shortening. A sufficient decrease in activation to stop muscle shortening can move the muscle to brake mode.
This view suggests that the engineering idealization of muscle as an arbitrary force source or as a selectable spring or viscoelastic element is less useful than viewing a muscle as a pair of more limited devices (a unidirectional motor and a brake) that switch automatically depending on the sign of the muscle velocity. Understanding the mechanochemistry of muscle allows us to go beyond Hill-type muscle models to better explain metabolic costs, force-velocity relationships, short range stiffness, force enhancement, and other muscle properties and nonlinearities, and make more accurate muscle models to simulate and predict behavior, as well as provide therapy, rehabilitation, and physical assistance. Understanding Nature's molecular motors can also help us build better robots.
The primary hypothesis is that muscle mechanochemistry supports a modal actuator, with two different modes. How muscle behaves mechanically and chemically is determined by the direction and amount of relative movement of the thin (actin) and thick (myosin) filaments, in addition to activation. Shortening muscle can be viewed primarily as a force source that uses metabolic energy and fatigues. Lengthening muscle largely acts as a brake or energy sink that does not use much metabolic energy and does not fatigue as rapidly. Muscle under all conditions including isometric has short range stiffness for stretches (but not releases) that does not use metabolic energy.
1) While shortening: crossbridges act as a unidirectional motor doing positive work and using energy (ATP)), or 2) While isometric or lengthening: crossbridges can hold a position against a load (acting like a spring with one end fastened) or resist a load lengthening the muscle (acting like a controllable variable brake [12]) without using energy (ATP). I believe mode 2 contributes to both short range stiffness and the force-velocity properties of muscle.
Viewing muscle as a modal actuator is really a statement about the statistics of crossbridge cycling. There are two kinds of crossbridge cycling, one using ATP and one not, and the type of crossbridge cycling is determined by the muscle velocity. crossbridge cycling will either be predominantly one type or another, and intermediate distributions are not likely. Given the mechanochemistry, it is difficult to see how muscle can act in any different way.
A second hypothesis is that the nervous system has to take this modal nature of muscle into account in generating appropriate commands.
A third hypothesis is that behavior has evolved or is learned so as to take advantage of the modal nature of muscle to minimize the use of ATP, and thus minimize metabolic cost.
A fourth hypothesis is that human adaptation to an exoskeleton will also take advantage of the modal nature of muscle to minimize the use of ATP, and thus minimize metabolic cost.
What is new here is the hypothesis that the distribution of crossbridges cycle-types has modes, and that the nervous system controls muscle by managing muscle velocity as well as force. I am emphasizing the macroscopic consequences of rapid detachment and reattachment without the use of ATP. I am not aware of proposals along these lines, but would not be surprised if they turned up as I further explore the rich body of work on muscle.
Another contribution is to update teaching about muscles. The coverage in textbooks does not make clear the implications of the mechanochemistry [34, 23, 2, 18].

We are starting by reviewing the "traditional" model of how muscle works. Here are some good books: Mechanism of Muscular Contraction by Jack A. Rall, Muscles, Reflexes, and Locomotion by Tom McMahon
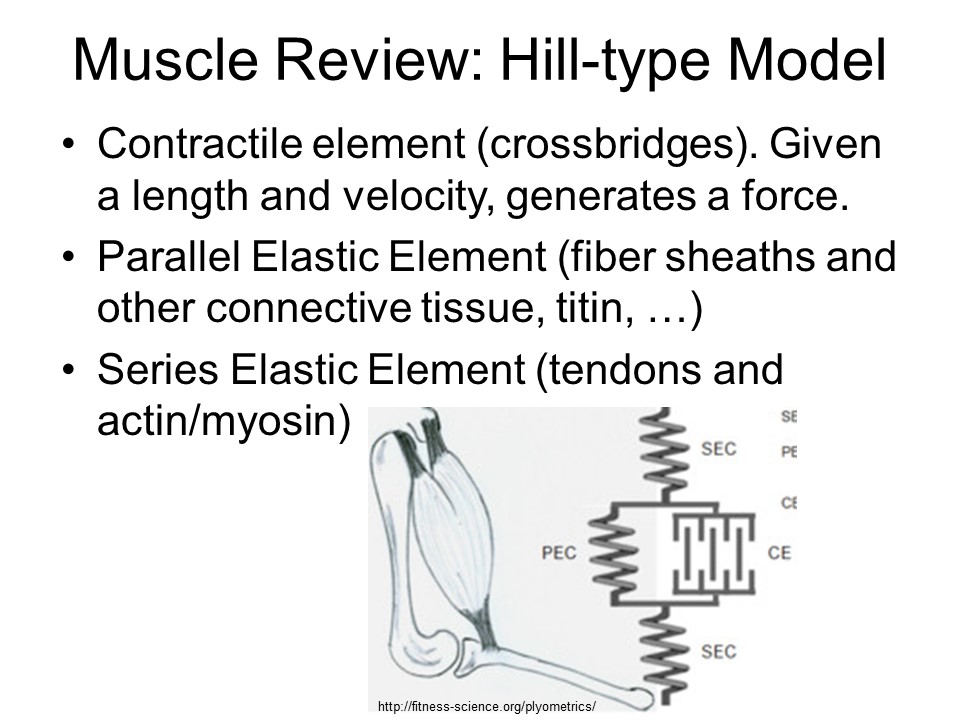
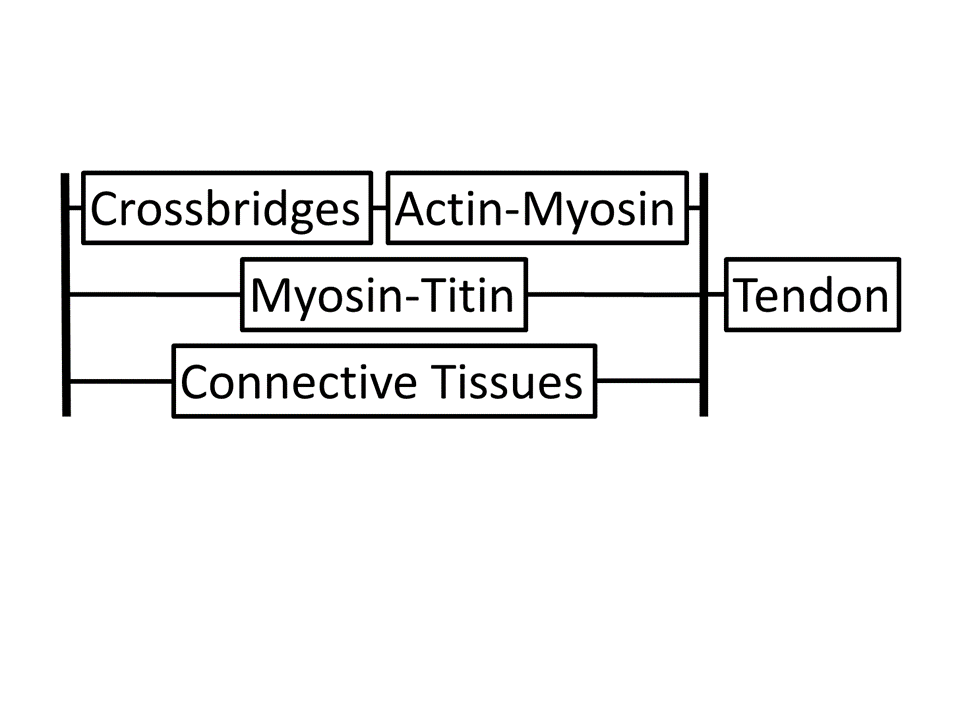
A whole-muscle view: Instead of the traditional picture showing a contractile element and showing separate springs as series elastic elements and parallel elastic elements, it is important to show the more complex mechanical arrangement and start to match mechanics with molecules. Force pathway 1: A contractile element is the conformational change of the myosin molecule, which is also elastic. Myosin filaments are also nonlinearly elastic as are the actin filaments. Multiple cross bridge connections means that the stress along the actin and myosin filaments changes at each crossbridge connection as some portion of the force is transferred across the crossbridge. Force pathway 2: Titin and myosin filaments form a second pathway for forces. If there is winding, the actin filaments will be involved. If there are Ca++ dependent changes to stiffness or equilibrium length of these chains, this adds further complexity to this force pathway, distinguishing it from other force pathways. Force pathway 3: Connective tissues provide another pathway for forces. I separate out the tendon as an overall series element to simplify the modeling. All of these pathways have nonlinear elasticity and viscosity (energy loss). A whole-muscle model would need to include all of these pathways. It is not clear what level of detail is required (lumped vs. filament-level).
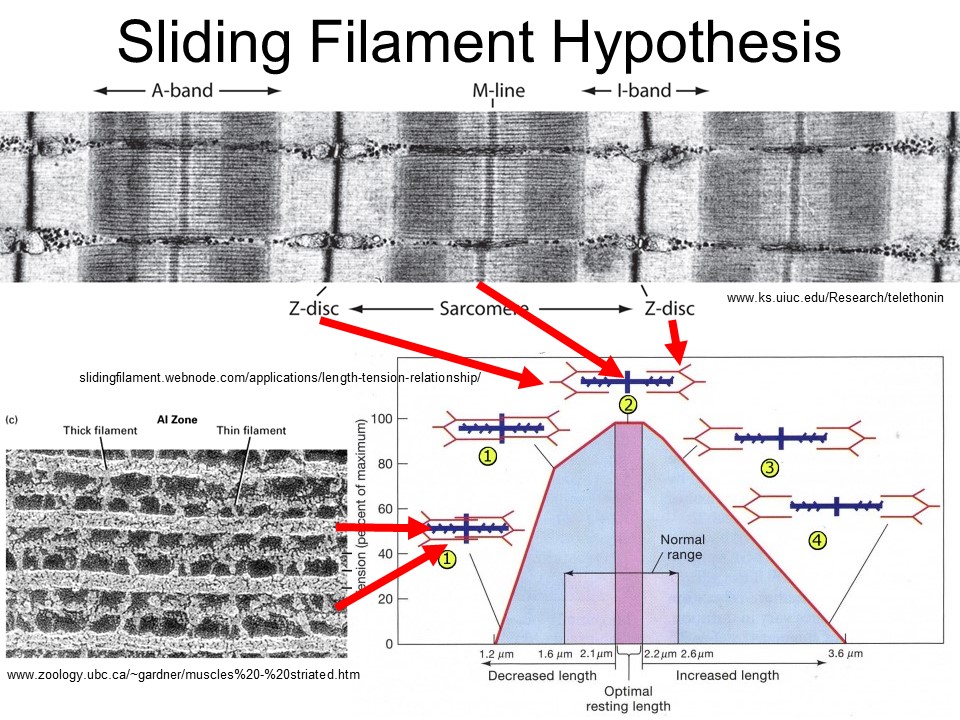
Actin (thin filaments) and myosin (thick filaments) slide relative to each other through the action of crossbridges.

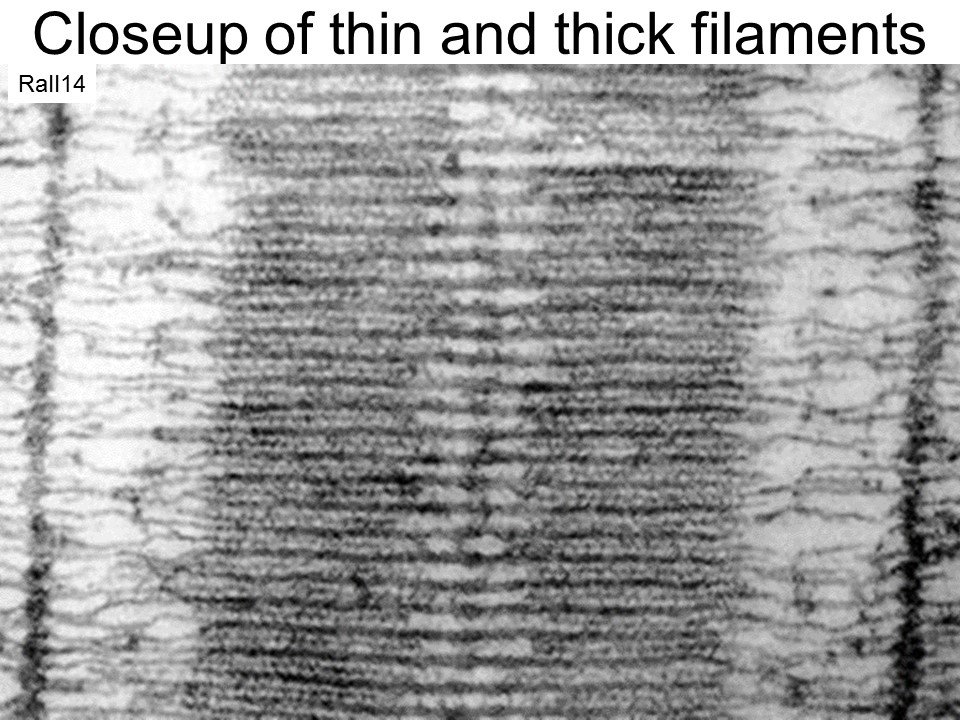
Titin, a protein which connects myosin to the Z line, is a hot topic which I will not discuss. It could attach or wind with the actin fiber when the muscle is activated. There are other muscle attachment proteins as well. Joke: One muscle protein is named "obscurin" because its function is obscure.

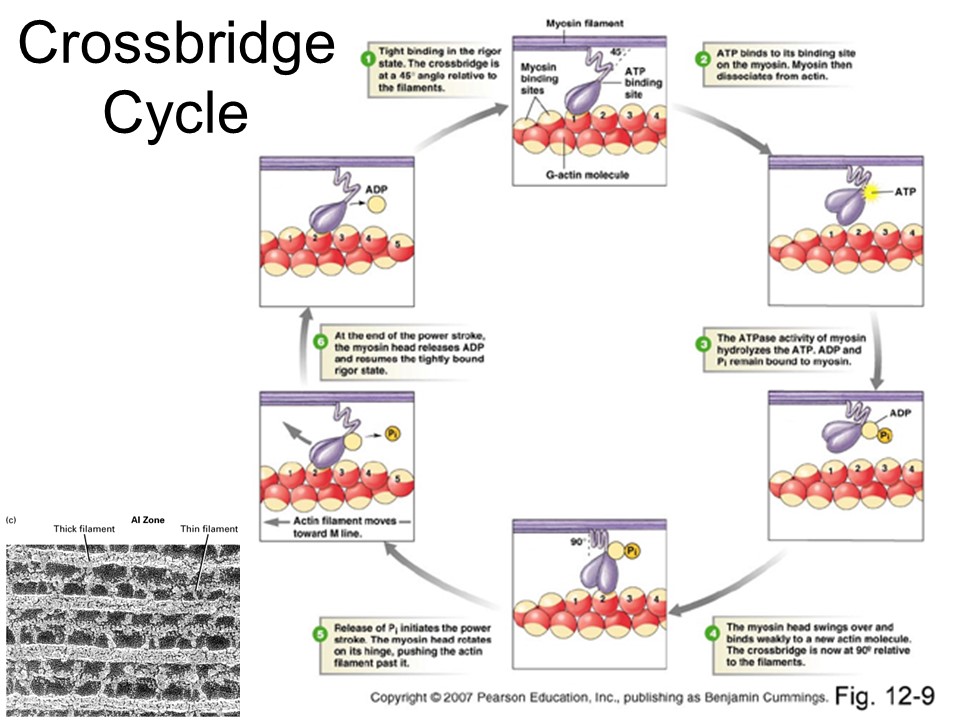
Here is my diagram of the traditional crossbridge cycle. I will add more pathways to it later.

In the next few slides I explain my previous view on how muscle works, and why the new view is such a big surprise to me. This paper is an example of "muscles are springs for control" school of thought.


Figure from Bizzi et al 1984.
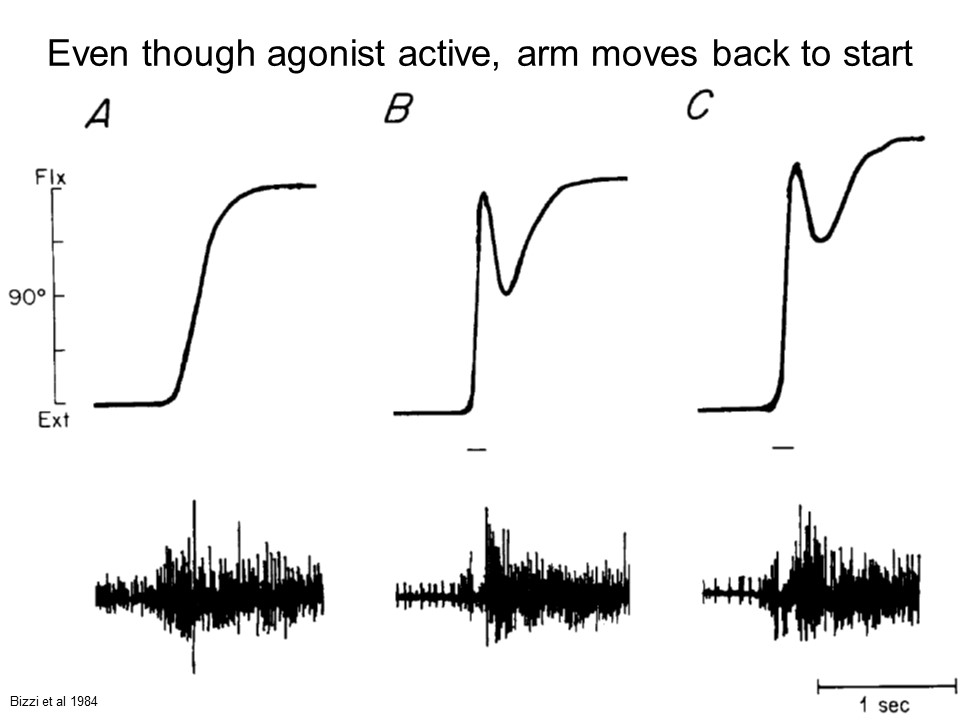

Figure from Bizzi et al 1984.

However, the view of muscle as a spring with fixed length-tension and force-velocity curves selected by activation is questionable. While tissues other than crossbridges may have fixed (but nonlinear) spring-like properties and generate the same force when returned to the same position and velocity, the macroscopic behavior of a set of crossbridges can be different depending on the history of the muscle activation and load, in addition to the current muscle length and velocity.
There is ample psychophysical and physiological evidence that muscles are at least twice as strong, use much less energy, are stiffer, and fatigue much less when producing a force during lengthening as compared to shortening.


Figure from Abbott et al 1952.
Figure from Elmer and LaStayo 2014. S. Elmer and P. LaStayo. Revisiting the positive aspects of negative work. Journal of Experimental Biology, 217:2434–2436, 2014.
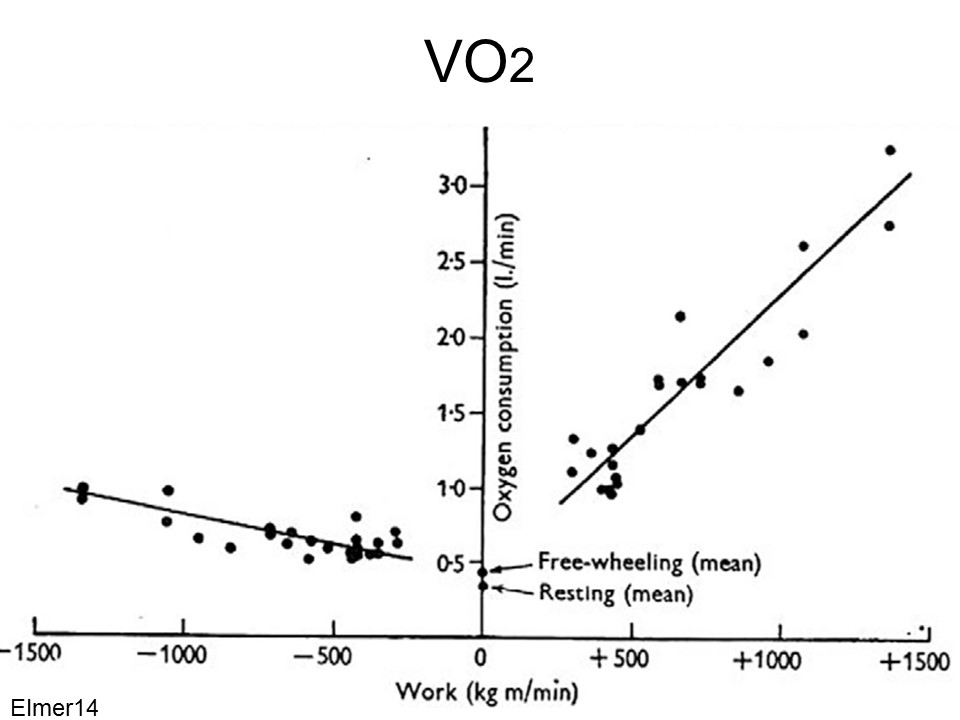
Figure from Bigland-Ritchie and Woods 1976.
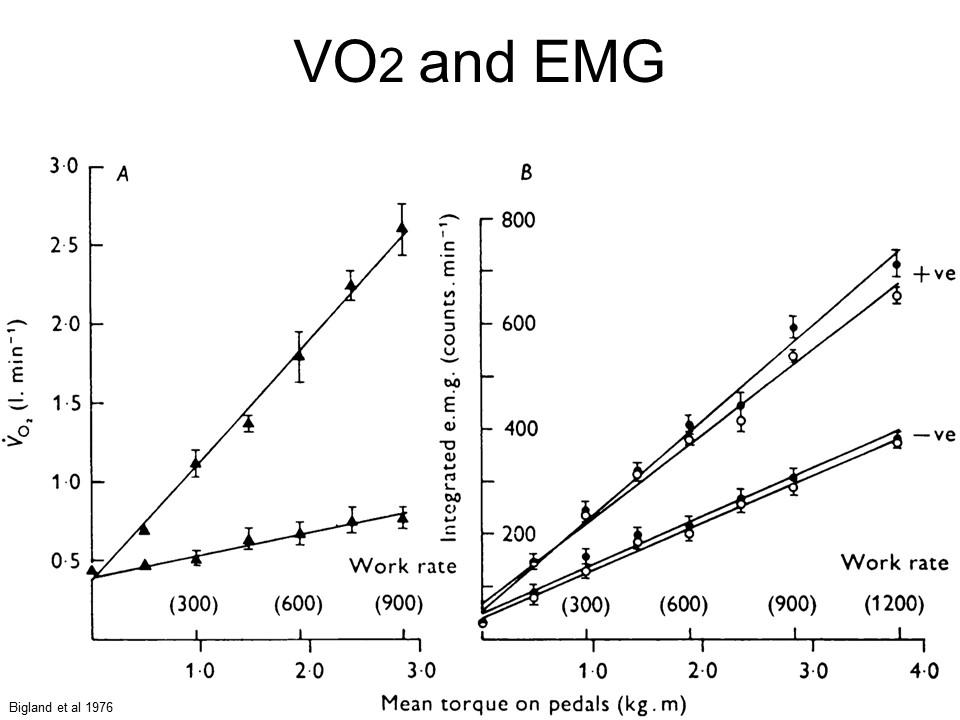
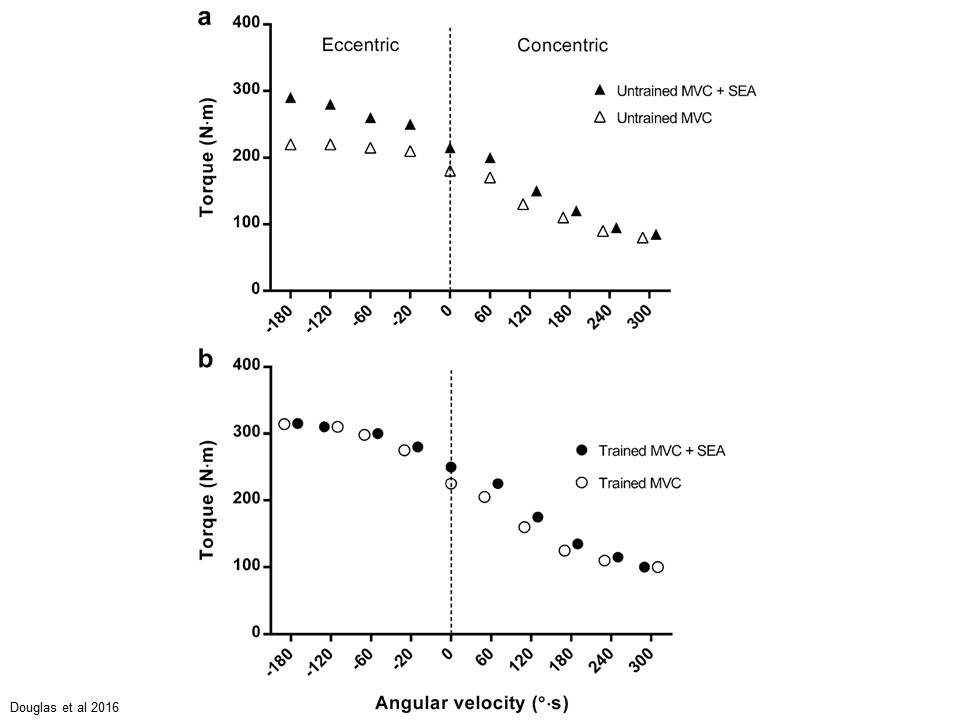
Figure from Douglas et al 2016.
In humans, the energy demand of eccentric and isometric work was negligible compared to concentric work, and only the concentric work significantly increased with force level, in a study using 31P magnetic resonance spectroscopy (MRS) to measure energy flow (the ratio of ATP-related phosphates Pi to PCr), Figure from Menard et al 1991. M. R. Menard, A. M. Penn, J. W. Lee, L. A. Dusik, and L. D. Hall. Relative metabolic efficiency of concentric and eccentric exercise determined by 31p magnetic resonance spectroscopy. Arch. Phys. Med. Rehabil., 72:976–983, 1991.
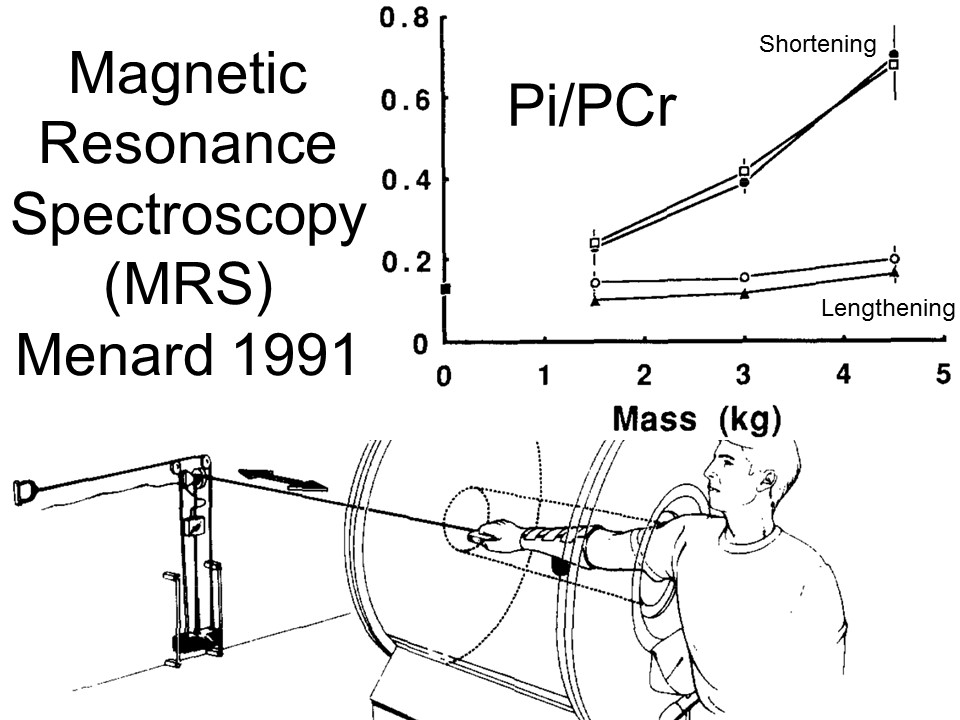
Figure from Curtin and Davies 1975.

In summary, it looks like muscle shortening is different from lengthening.
 \
\
Now we will describe hypotheses about the underlying molecular mechanisms that make muscle shortening and lengthening different. Here is the traditional crossbridge cycle.
The previous slide makes it look like only one chemical pathway happens. In actuality all possible transitions happen, just with different rate constants.

Rate constants for some pathways are affected by the conformation of the molecules involved. Stretching the crossbridge elastic element clearly changes the conformation of the myosin (and possibly actin) molecule(s).

It is hypothesized that the release rate of ADP by the myosin head is reduced by crossbridge stretch (strain). In muscle shortening, eventually each crossbridge will shorten as the filaments move relative to each other, and the strain will be low enough that ATP can bind and the crossbridge can detach. In muscle lengthening, crossbridge strain increases, and the rate of crossbridge detachment becomes lower.
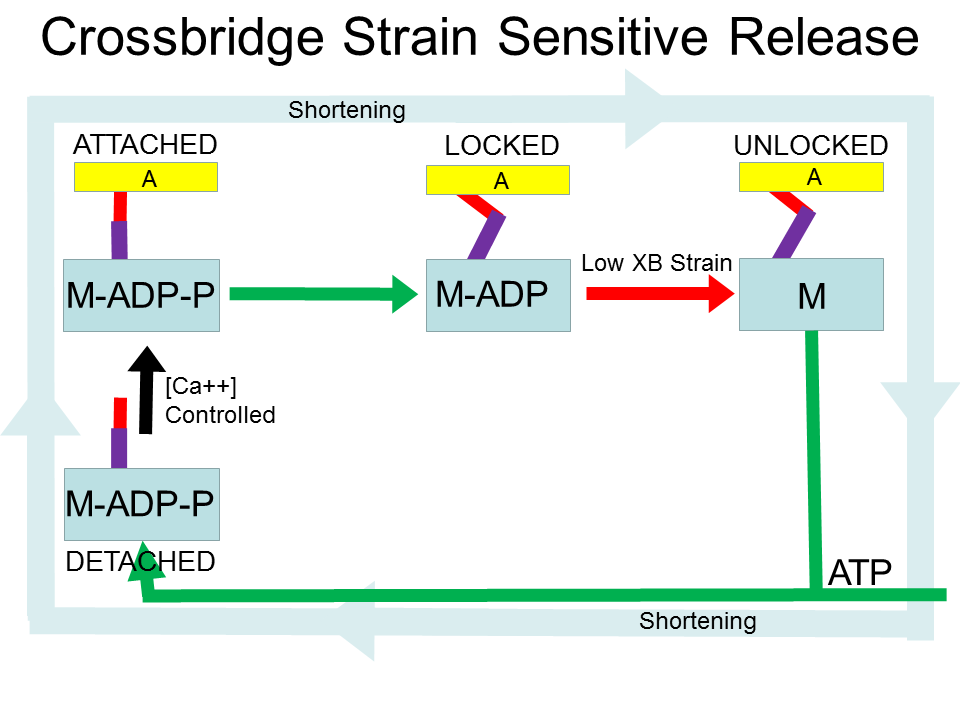
Here are examples of strain dependent mechanisms. The Chinese Finger Trap can be released by allowing it to shorten and increase its diameter. Cam and jam cleats and friction locks use conformational change of the cleat and/or the rope to lock onto the rope. They are easier to release if the rope is slack. Wrapping a rope around a round object (winch, capstan, dock pillar, ...) allows a sailor to gradually release the rope, aided by the friction between the rope and the round object. The amount of friction depends on the load on the rope and thus the stretch (strain) of the rope.

The Huxley 1957 model included this hypothesis, in that the detachment rate (g) is much larger for negative crossbridge strains. "The rate constant for detachment is small as long as the cross-bridge is exerting positive tension but becomes large as soon as shortening has brought the cross-bridge past the position where the force it exerts is zero." [20]

The crossbridge cannot remain attached as the muscle lengthens. It is hypothesized that it eventually detaches, but without needing ATP to do so. Caremani et al 2015. The crossbridge is still in the "cocked" or "energized" state.

It is also hypothesized that the "energized" crossbrige rapidly reattches to the actin molecule at a binding site where the crossbridge is much less stretched. Huxley in 2000 reviewed evidence for crossbridge "jumping" without the use of ATP [20]. Caremani et al 2015.
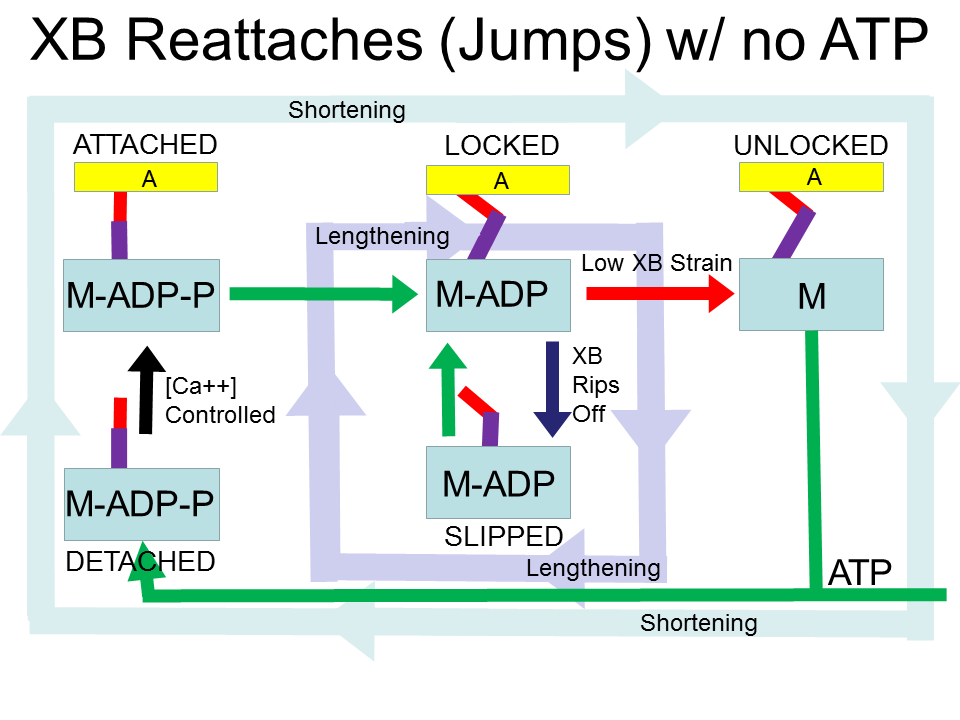
Caption: The myosin cycle. Green arrows show rapid transitions. Arrows with other colors show proposed controlled transitions. The black arrow shows where calcium ions (Ca++) control the attachment of myosin heads (M-ADP-P, blue block) to the actin (A, yellow block) thin filaments. The red arrow shows where low crossbridge (XB) strain permits transitions to the UNLOCKED state. The blue arrow indicates where crossbridge strain that exceeds a mechanical limit causes detachment and slipping. The faint blue cycle shows the traditional dominant pathway when the muscle is shortening. The faint purple cycle shows the proposed dominant pathway when the muscle is lenthening, a cycle of detachment and attachment that leads to a viscous-like resistance to changes in muscle length, without using ATP. In the loaded isometric case the crossbridges remain in the locked state. The purple line is the lever arm of the myosin head, and the red line is a hypothesized elastic element. The details of where the elasticity actually resides in the myosin molecule are not crucial to the theory.
Let's walk through this starting with the DETACHED state. Calcium ions control the transition to the ATTACHED state. Note that the initial stretch of the crossbridge is zero. Right away (the likely and fast pathway) a phosphate P is released and the myosin molecule reconfigures (power stroke) to stretch the crossbridge and add a little pulling force (a few pico-Newtons) to the muscle.
It is very important to keep in mind that a single crossbridge does not determine the relative motion of the actin and myosin filaments. The length of the crossbridges is determined by the state of all of the crossbridges, the overall muscle length, and forces in and stiffness of the various filaments (actin, myosin, titin, ...) and other passive tissues involved. Since mass is moved, this entire system has dynamics, so the past states of the system also matter. Furthermore, muscle and tissue are not rigid bodies, and the distribution of motion and mass is quite complex. The compliance of crossbridges allows the crossbridges to stretch as necessary to accommodate some of this movement.
Now comes the interesting part. In order to continue the cycle, a myosin molecule must release the bound ADP molecule to be able to detach from the actin filament The probability of this transition is reduced by crossbridge stretch (recently shown in single skeletal muscle myosin II actin-myosin bonds by [7, 8]; [36, 16, 29] provide reviews). If the crossbridge is stretched (contributing to the pulling force of the muscle), it is not likely to finish later stage(s) of the power stroke, release the ADP molecule, and detach. This is a form of “catch bond” (https://en.wikipedia.org/wiki/ Catch_bond) [17]. The crossbridge is LOCKED, acting like a Chinese Finger Trap which locks on to a finger when pulled (Figure 2), and contributing to the local spring-like behavior of muscle. Variants of this property have been found in non-skeletal muscle myosins as well [36, 16, 29, 21, 40]. There are hints that this property may be more pronounced in slow isoforms of myosin, and thus may be more important in slow-twitch motor units [6]. I believe the locked crossbridges strongly contribute to the overall short range stiffness of muscle.
A key element of muscle energy efficiency is the ability to cycle the crossbridges while resisting a pulling load without using ATP. If a crossbridge is stretched too far while in the LOCKED state, the crossbridge slips by detaching to the SLIPPED state and then quickly re-attaching. The crossbridge is back in the LOCKED state at a new actin binding site where it is stretched less. I call this a "jump". It could also be called a "slip-and-catch". The myosin "lever arm" remains in the "pulled" conformation. If the muscle continues to lengthen, this cycle continues, contributing a resisting force and turning externally provided work into heat [8]. This cycle is a source of friction/viscosity (resistance and energy loss due to velocity) of muscle (often modeled using an overall force-velocity curve). The amount of resistance and energy loss can be modulated by the amount of crossbridges in the LOCKED state, which is throttled by the concentration of Ca++ ions.
If the muscle needs to actively shorten, increasing the Ca++ concentration recruits more crossbridges and reduces the stretch of each individual crossbridge. If a crossbridge is relaxed, it is more likely to move to the UNLOCKED state, and quickly moves to the DETACHED state (using ATP) through various possible pathways, the details of which are not crucial to the proposed theory. As the elastic element shortens to the equilibrium length, the rate of releasing ADP and exiting the LOCKED state increases by an order of magnitude [8].
Although the distribution of the type of crossbridge cycles is determined by the muscle velocity, a command to a muscle generated by the nervous system can cause that muscle to hold its length or resist a pull (brake mode) by stopping the muscle lengthening. Additional activation can cause the muscle to switch modes and shorten by causing the muscle to start shortening. A decrease in activation can return muscle to the brake mode. To control modes, the nervous system can control the sign of the muscle velocity.
The above diagram shows a minimum set of molecular states that implement the proposed theory with the proposed multiple control points. There are proposals for finer divisions of all parts of this cycle [29]. Additional states can be accommodated in this theory, as long as ATP-fueled detachment requires low strain in the crossbridge, and rapid crossbridge cycling can occur driven by large crossbridge strains without the use of ATP.
There are several control steps in the mechanochemical actin-myosin cycle. 1) The concentration of calcium ions (Ca++) controls the rate of attachment of crossbridges (with the crossbridge unstretched at attachment). 2) The amount of stretch (strain) in the crossbridge controls the probability the crossbridge will detach. Normal detachment using ATP requires low strain in the crossbridge. High strain changes the conformation of the crossbridge so that the myosin head is locked to the actin filament, and cannot detach. 3) In any attached state, if the crossbridge is stretched too far (strain > mechanical limit), the crossbridge will detach and then quickly reattach with the myosin "lever arm" in the same conformation and the elastic element near equilibrium, with no consumption of ATP.
Another hypothesis is that a stretched crossbridge has a cooperative effect on the probability the 2nd myosin head will also attach, making detachment of that myosin molecule less likely.

Let's consider arguments against the idea that muscle lengthening is different from shortening, or that it matters.

It is true that eccentric exercise can damage your muscles. You will be sore 1-2 days after you increase the amount of eccentric exercise you do. You will eventually recover. I hypothesize eccentric work that is part of your daily routine does not cause muscle damage.


How much eccentric work do we do daily? I argue that wrist velocity profiles are roughly symmetric, and we do about as much deceleration as acceleration. Assuming there is little physical damping in the arm, deceleration forces are about the same as acceleration forces, on average. For example, the amount of upward movements is balanced by the amount of downward movements. The characteristics of upward and downward movements are the same, Humans don't change the accelerations or decelerations in this case.


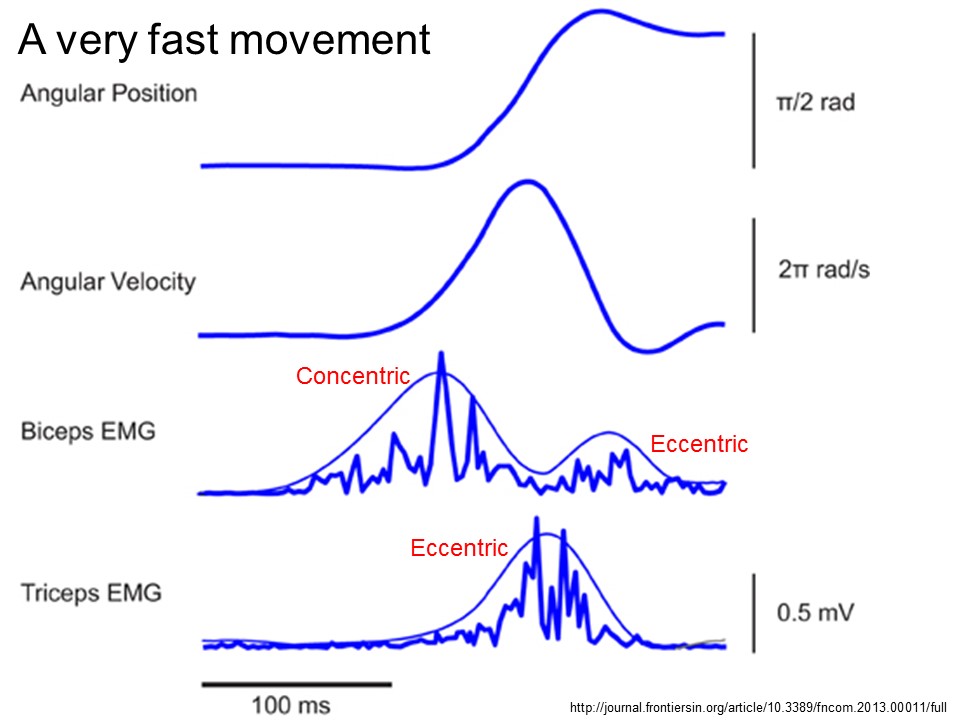
Now let's look at muscle activation (electromyographic activity (EMG)) during these types of movements. We see muscle activity during deceleration, which indicates muscles are active while lengthening. Figure from Gribble et al 2003

Figure from Chiovetto et al 2013
Looking at muscle fiber lengths during walking, we see many examples of active muscles lengthening and then remaining the same length (isometric). A hypothesis is that lengthening increases the number of cross bridges and thus the strength of the isometric muscle.
Muscles acting as brakes are an important part of legged locomotion [12], and may also play an important role in whole-body posture, particularly when compensating for gravitational forces. In the case of muscles, the use of brakes costs no energy, and uses no additional material, space, or weight. Delp's group has published estimates of muscle fiber length during gait [1]. The following figure shows many examples of muscle activation during isometric and lengthening phases. Biewener proposes that distal limb muscles are specialized to act like brakes and heavily geared motors, while larger limb proximal muscles are specialized to do positive work over larger distances [3].
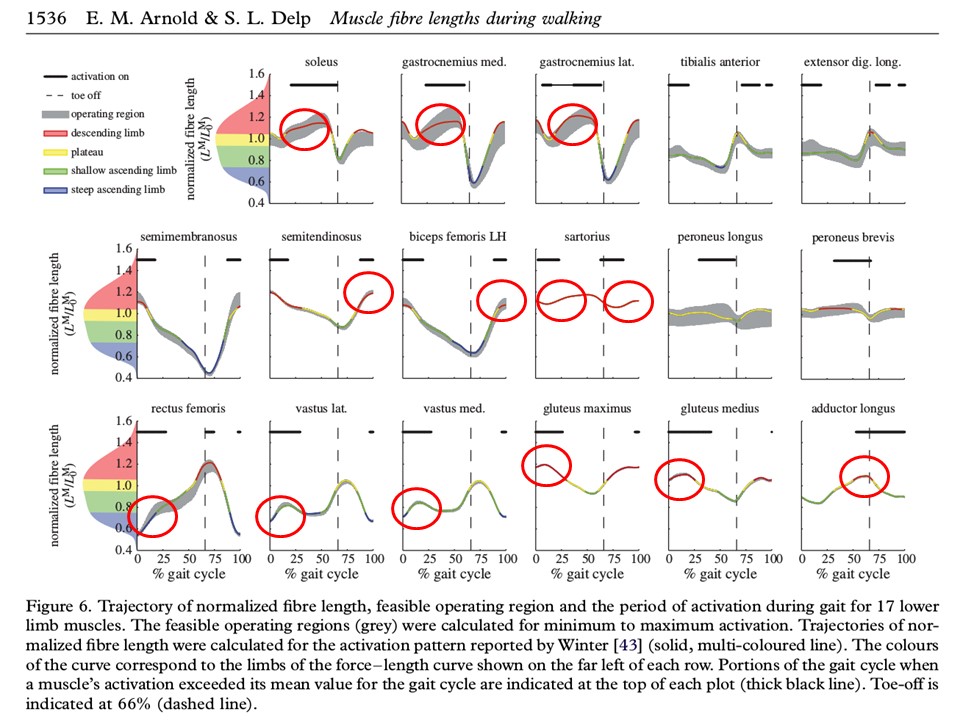
Figure from McGowan et al 2010
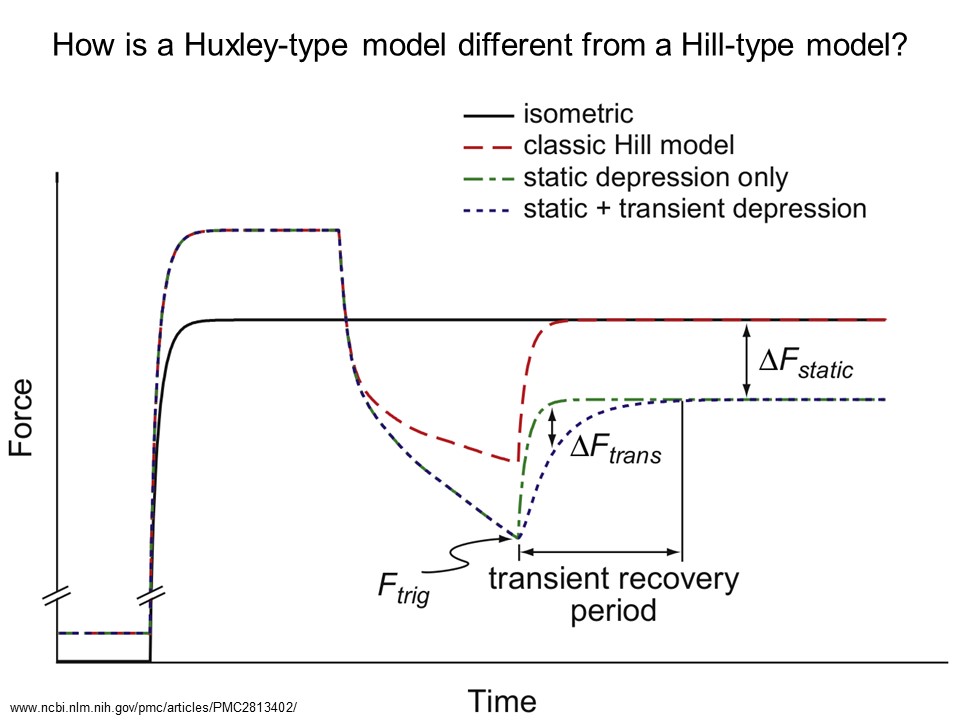
Figure from McGowan et al 2010

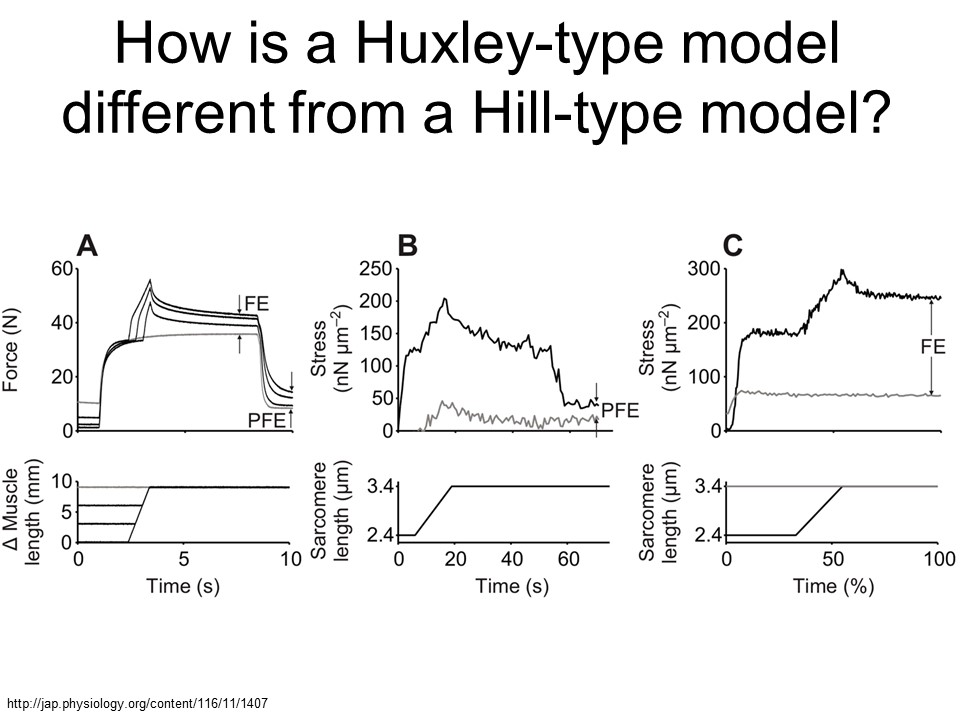
Figure from Herzog 2014
The shaded portions below are measured human elbow stiffnesses. The lines on the left that do not match well are based on the slope of the length-tension curves. The lines on the right that fit better are based on estimates of short range stiffness. Figure from Hu et al 2011

Another paper just came out with similar conclusions: "Contribution of muscle short-range stiffness to initial changes in joint kinetics and kinematics during perturbations to standing balance: a simulation study", Friedl De Groote, Jessica L. Allen, and Lena H. Ting, Journal of Biomechanics, 2017


Current torque-controlled humanoid robots are now successfully walking largely because torque control is finally good enough: achieved with high quality expensive and delicate sensors, and an order of magnitude less feedback latency than biological systems. However, it is notoriously difficult to damp out oscillations in torque controlled robots, and especially robots with series elastic actuators (for example, MIT's M2 NEED URL HERE). Motors with low gear ratios use a lot of energy to hold a position or resist a push. Electric motors produce torque that is proportional to current, and the power used is the current multiplied by the power supply voltage. Hydraulic motors can hold a position with no energy cost by closing valves. However, resisting a pull while moving has an energy cost of the fluid flow multiplied by the supply pressure. In addition, force-controlled hydraulic systems used on humanoid robots typically have significant continual energy loss through internal leakage, which is necessary to keep friction in pistons low. It is clear that robotics has been pursuing a mistaken design paradigm. Because our abstractions about muscle were wrong, we missed a better design approach: a set of actuators at each joint: inaccurate but repeatable unidirectional motors and physical brakes, similar to the actuation of automobiles.
This view of muscle as a set of distinct actuators contrasts with the typical view in humanoid robotics that actuators should be high quality torque sources, and should be able to track arbitrary desired trajectories. I am advocating that behavior should be generated by selecting one of the available modes for each muscle at each moment. It turns out that mode selection is largely automatic. If the muscle is shortening, it shortens more (in the motor mode). Otherwise, the muscle acts like a brake with a controllable maximum friction level. To switch from brake mode to motor mode, an excess amount of activation (or a decrease in the external load) to initiate shortening is required. This is how a one dimensional command can control an actuator with two modes. Maybe robots should work the same way.
A better understanding of how muscles work will lead to higher performance robots. One day we may be able to engineer molecular motors. In the meantime, we can focus on simulating the muscle properties and behavior described above with a combination of simple inexpensive actuators: unidirectional motors (no backlash issues) and variable brakes (greatly reducing stability issues and the need for high quality torque and velocity feedback).

This robot from the early 1990s had physical springs and dampers in its hands. Movie on Youtube. A paper.

Engineering examples of multiple actuators for the same DOF:
Need to list papers.


I am looking for muscle models I could modify to test this hypothesis, or potential collaborators who would be interested in exploring this hypothesis.
More to come on hidden assumptions I made and unresolved questions:
I assume long stretches of binding sites on actin are available. If there are not sufficient streches of actin binding sites, the ripped off myosin head cannot reattach.
Is Ca++ necessary to maintain actin-myosin bond, or only to create the bond? - no, rigor is maintained without Ca++ (Bayshaw). Is Ca++ necessary to maintain reattachment process while lenthening? Does the brake mode persist after activation stops?
There is some weird stuff on measurements of short range stiffness in the human wrist where it is time (30ms) rather than distance based [11]. If it is really time based, that needs some re-thinking about what makes crossbridges detach and how long they can physically be.
It may be the case that short range stiffness is actually generated by other molecules such as titin, which seems to be a source of Ca++ modulated passive stiffness.
There is an issue that non-uniform sarcomere length distributions generate effects like force enhancement [5].
Other strain based mechanisms:
Revisit intrafiber sarcomere length stability. Sarcomeres in the same fiber all have roughtly the same Ca++ activation (we assume). They have local positive stiffness, but globally some can fully shorten while others fully lengthen. What prevents this? Passive stiffness of actin, myosin, or titin? Passive stiffness of the rest of the fiber tissue? Neighboring fiber stiffness?
What About Winding Filaments? I see this as an orthogonal or complementary issue. Winding filaments still have actin-myosin bonds with the proposed properties, so the behavior should be similar. However, winding filaments could increase actin-myosin-titin tension and stiffness in addition to cross bridge activity.
What About Ca++ Dependent Passive Stiffness (Titin)? I see this as an orthogonal or complementary issue, closely related to the winding filament hypothesis. Intramuscular passive stiffness should increase with activation, and would augment stiffness due to crossbridges. It may be the case that short range stiffness is also generated by other molecules such as titin, which seems to be a source of Ca++ modulated passive stiffness. The collapse of short range stiffness could be due to the passive unwinding of filaments.
What about co-contraction?
How do slow twitch motor units keep up with fast twitch motor units when both are recruited? How does slow myosin keep up with fast myosin when both are recruited?
Does this explain why biological systems are usually well damped?
What is going on when muscles are activated before impact?
Google "Engineered skeletal muscle" to get pointers to people trying to grow or reconstruct muscle for engineering applications.
Track down hypothesis that single ATP powers 5-6 detachments. See Rall 14 counterargument about duty cycle.
Is this true?: Brakes need to be large (and typically heavy) to generate sufficient force and dissipate the resulting heat, since there are limits on the friction coefficient that a practical controllable variable brake can have.